Palynological investigations of the James Ross Island ...
Feldmann, R.M., and W.J. Zinsmeister. 1984. Fossil crabs (Decapoda: Brachyura) from the La Meseta Formation (Eocene) of Antarctica: Paleoecological and biogeographic implications. Journal of Paleontology, 58(4), 1046-1061. Macellari, C.E. 1986. Late Campanian-Maastrichtian ammonite fauna from Seymour Island (Antarctic Peninsula). Journal of Paleontology, Vol. 60, (The Paleontological Society Memoir 18, Part 2.) Sadler, P.M. In press. Geometry and stratification of Paleogene and latest Cretaceous units on Seymour Island, northern Antarctic Peninsula. In R.M. Feldmann and M.O. Woodburne, (Eds.), Geology and paleontology of Seymour island, Antarctica. (Geological Society of America Memoir 169.) Tshudy, D., and R.M. Feldmann. In press. Macrurous decapod crustaceans, and their epibionts, from the Lopez de Bertodano Formation (Late Cretaceous), Seymour Island, Antarctica. In R.M. Feldmann and M . O. Woodburne, (Eds.), Geology and paleontology of Seymour Island, Antarctica. (Geological Society of America Memoir 169.)
-j
Figure 2. Hoploparia stokesi Weller. Latex cast of a specimen from the Lopez de Bertodano Formation, preserved as a molted, but complete, carapace and abdomen. (Bar scale equals 1 centimeter.)
Palynological investigations of the James Ross Island basin, Antarctica ROSEMARY
A. ASKIN
Department of Earth Sciences University of California Riverside, California 92521
During December 1986 and January 1987, additional sampling of Campanian to Eocene sediments was carried out for palynological study in the James Ross Island basin (figure), northeastern Antarctic Peninsula. The 300 samples collected are mainly from Seymour Island and cover certain intervals of interest in more detail than was previously available. Data from these samples will refine and extend the palynomorph zonation proposed for the upper Campanian to Paleocene part of the succession (Askin in press). Parts of the upper Lopez de Bertodano Formation, including the glauconitic interval that spans the Cretaceous/Tertiary boundary, and parts of the Eocene La Meseta Formation were sampled. Short helicopter-supported visits also were made to three nearby islands to collect from ?Campanian sections. The three sampled localities are Cockburn Island, Humps Island, and Ula Point on eastern James Ross Island (figure). Fossil palynomorphs are ubiquitous in the Seymour and nearby island sedimentary succession and occur, often abundantly, in all 935 samples processed to date. It is expected that the newly collected samples also will be productive, bringing the samples available for this study to a total of over 1,200. The present phase of this project includes taxonomic description of nonmarine (spores, pollen) and marine (dinoflagellate 1987 REVIEW
cysts, acritarchs) palynomorphs and documentation of their distribution patterns. The dinocyst assemblages have proved particularly useful for biostratigraphy (Askin in press), and the land plant-derived spores and pollen should add useful information on climatic evolution of the area. Unlike terrestrial sediments, which can contain palynomorphs of relatively local derivation, the mainly nearshore marine sediments of Seymour Island contain spores and pollen transported from a wide range of terrestrial habitats and microclimates. Thus these as-
Vega Island
(J c::;
AAfarctica
Naze
640
Island
9
5 10 15 20
VUIO Point
I)
I James Ross Island
(
Km
Cockburn
Island
Hamilton
Seymour Island EPateogene sediments []
Cretaceous sediments
Snow Hill
Island
Locality map for the James Ross Island basin. ("Km" denotes "kilometer?")
13
semblages will provide a broad view of the prevailing paleoclimatic conditions. The Cretaceous cryptogam (ferns and lower plants) spores of Seymour Island (Askin in preparation) include an overwhelming abundance of Stereisporites antiquasporites and Laevigatosporites ovatus. These two species, plus fewer numbers of S. regiuln, usually total over 75 percent of the spore component. The abundance of Stereisporites (the spores of Sphagnum moss), along with some Azolla sp. (water fern), suggests moist conditions with ample standing water, consistent with humid temperate paleoclimates indicated both by the palynomorph assemblages (dominated by podocarpaceous conifer pollen) and by other lines of evidence (e.g., fossil wood; Francis 1986). Alternatively, hydrodynamic properties of the small Stereisporites and Laevigatosporites spores may have contributed to their concentration. Analysis of all the acid-insoluble organic components of the sediments (palynomorphs, plus cuticle, amorphous and algal matter, and woody debris) should help
answer such questions, by providing inferred data on proximity to shoreline, higher vs. lower energy environments, and so on. I thank F.C. Barbis and T.R. Kelley for 1986-1987 collection of samples, and the Captain and crew of USCGC Glacier for logistic support. This research is supported by National Science Foundation grant DPP 83-14186.
A study of the internal annual growth lines of the Late Eocene mollusk
profiles from the planktonic forams in the oceans surrounding Antarctica (Kennett 1977) show that the sea-surface temperatures in the southern oceans during the Late Eocene were not significantly different from the sea-surface temperatures off the coast of Chile today. It is believed that the difference in patterns is due to low levels of plankton productivity in the winter months at the high latitudes when the daily levels of sunlight are not high enough to maintain significant photosynthetic levels. In a recent study, Tilzer and Dubinsky (1987) found that respiration rates in phytoplankton are temperature-dependent, with respiration rates decreasing with temperature. Plants lose carbon during respiration so the mass balance of the population is controlled by the respiration rate. The phytoplankton are therefore able to maintain a positive mass balance during the long antarctic winter because they are able to conserve carbon and stored energy at such low temperatures. If the temperature was raised even a few degrees, then the respiration rates increased enough that it was impossible for the phytoplankton to maintain this positive mass balance. During the Late Eocene, the average surface seawater temperatures in the southern oceans were approximately 9° to 12°C higher than they are today (Kennett 1977). According to Tilzer and Dubinsky (1987), the phytoplankton population would have been greatly reduced during the winter months. There-
Eurhomalea antarctica
BRENDA L. DEXTRAZE and WILLIAM J. ZINSMEISTER
Department of Earth and Atmospheric Sciences Purdue University West Lafayette, Indiana 47907
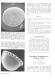
During the last decade, much work has been done using the internal growth increments annually recorded in bivalve shells. These growth increments are believed to record the slowing down or cessation of growth of the bivalve during the cold winter months (Panella and MacClintock 1968). These growth lines are analogous to tree rings and are very useful environmental indicators. They record the ambient seawater temperatures (Urey et al. 1951) as well as the geochemical history of the seawater in which the bivalve lived (Eisma, Mook, and Das 1976). In this study, the internal annual growth lines of two Late Eocene antarctic mollusks (Eurhomalea antarctica and E. newtoni) from Seymour Island (64.5°S) were compared to the growth lines from a Recent Eurhomalea species (E. lenticularis) from two South American localities, Chile at 34.5°S and Patagonia at 55°S. No significant difference can be seen between the growth increment patterns of the Chilean and Patagonian specimens. These individuals of Eurhomalea show a long period of growth broken by thin annual increments. It is believed that this pattern reflects fast growth during most of the year and slow growth during the winter months (figure). Eocene shells from Seymour Island show a different type of growth increment pattern: an extremely short period of growth followed by a thick winter line. In most cases, temperature regime is considered to be the primary cause of growth increment patterns in bivalves. In this case it does not appear that the temperature regime dictated the growth increment patterns. This is because O temperature 14
References
Askin, R. A. In press. Campanian to Paleocene palynological succession of Seymour and adjacent islands, northeastern Antarctic Peninsula. In R.M. Feldmann and M.O. Woodburne (Eds.), Geology and paleontology of Seymour Island, Antarctic Peninsula. (Geological Society of America Memoir 169.) Askin, R.A. In preparation. Cryptogam spores from the upper Campanian and Maastrichtian of Seymour Island, Antarctica. Francis, J.E. 1986. Growth rings in Cretaceous and Tertiary wood from Antarctica and their palaeoclimatic implications. Palaeontology, 29, 665-686.
Thin section of Eurhomalea lenticularis from central Chile, 34.5°S. ANTARCTIC JOURNAL